
Недавно две группы ученых убедительно доказали, что одной из причин старения и болезни Паркинсона являются дефектные митохондрии, быстро размножающиеся в нейронах головного мозга и приводящие к их дегенерации. Дефектными они становятся в результате соматической мутации - удаления участка митохондриальной ДНК. Однако остается неясным, почему с возрастом дефектные митохондрии появляются в определенных нейронах головного мозга.
Пока нет общепринятого мнения о причинах старения. По одной из гипотез, старение и смерть индивидуумов, не способных более к размножению, выгодно для вида, поскольку экономятся ресурсы (пища, территория), необходимые для молодых, более полезных для существования всего вида особей. Хотя не известно точно, сколько генов задействовано в этой программе старения и как они взаимодействуют, но, детально изучив их, можно попытаться заблокировать механизм старения.
Согласно же другой гипотезе, старение - результат действия сотен или даже тысяч вредных генетических признаков, которые начинают проявляться лишь по окончании репродуктивного периода. Мы будем называть их DILL-признаки (DILL - от англ. Deleterious In Late Life ), или DILL-аллели.
Почему эти DILL-признаки накапливались в течение эволюции? Выделяют три возможные причины (Cortopassi 2002). Во-первых, численность популяции на протяжении большей части эволюции человека была крайне низкой (примерно 10 000), в связи с чем случайные процессы сильно влияли на эволюцию генов человека и приводили к закреплению множества вредных аллелей. Недавнее резкое увеличение численности популяции еще не отразилось на эволюции генов человека.
Во-вторых, мутация, проявляющаяся лишь после репродуктивного периода, очень слабо влияет на приспособленность организма (число детей) и поэтому может распространиться в популяции случайным образом, без влияния естественного отбора.
И в-третьих, даже если предположить, что DILL-мутации как-то влияют на приспособленность (например, мудрые советы пожилых людей обеспечивают лучшую выживаемость их детей и внуков), естественный отбор не мог элиминировать эти мутации, поскольку в древней человеческой популяции средняя продолжительность жизни была низкой. За 5 миллионов лет могло накопиться множество мутаций, вызывающих ухудшение слуха в 50 лет, ослабление памяти в 55, болезнь Паркинсона в 60, дряблость кожи в 65... Все они находились в полной безвестности, пока примерно 10 000 лет назад человек не занялся земледелием, продолжительность его жизни увеличилась, и все эти DILL-мутации стали проявляться. Начиная с этого момента человек стал умирать преимущественно от внутренних причин
Однако гены накопили DILL-мутации неравномерно, поэтому и потенциальная вредность от этих генов в старости будет разная. Например, митохондриальные гены (те которые содержатся в митохондрии, наследуемой через цитоплазму яйцеклетки от матери) должны содержать много DILL-мутаций. Это объясняется тем, что в связи с гаплоидностью, цитоплазматическим наследованием и отсутствием рекомбинации митохондриальные гены обладают низкой эффективной численностью, поэтому на их эволюцию сильно влияют случайные факторы и слабо влияет естественный отбор. В результате дрейфа генов в малочисленной популяции митохондриальных генов могут фиксироваться неоптимальные аллели, содержащие вредные мутации.
Кроме того, митохондрии обладают повышенным темпом мутирования, что увеличивает вероятность появления новой соматической мутации в течение жизни человека. Но, поскольку в каждой клетке содержатся тысячи митохондриальных геномов (в среднем около 2000), они создают генетическую избыточность, и один появившийся мутант, сосуществуя с нормальными митохондриальными геномами, не может сильно повлиять на приспособленность клетки. Дальнейшая судьба мутантного генома зависит от соотношения сил на внутриклеточном и межклеточном уровнях (Taylor 2002).
Внутри клетки митохондриальные геномы конкурируют друг с другом. Геномы, которые быстрее реплицируются, увеличивают свою численность внутри клетки и вытесняют медленно-размножающихся конкурентов. Если у одного генома произошла крупная делеция (вырезание участка ДНК), то он становится нефункциональным (так как производится меньше АТФ - энергетической молекулы, необходимой для ускорения большинства биологических реакций), но зато коротким и быстро размножающимся - и такой эгоистичный геном победит во внутриклеточной борьбе.
Однако межклеточный уровень конкуренции ставит всё на свои места: клетка, в которой мутантные митохондриальные геномы достигли высокой концентрации, менее жизнеспособна и медленнее делится по сравнению с клеткой с нормальными митохондриальными геномами, и мутантные геномы элиминируются из организма вместе со своей клеткой.
Важно отметить, что порог проявления патогенного эффекта мутантного генома варьирует в зависимости от энергетической потребности клетки. Так, ткани, сильно зависящие от митохондриального метаболизма (мышцы, сердце и нервная система) более чувствительны к накоплению мутантных геномов и, значит, имеют самый низкий порог.
Помимо своей высокой энергетической емкости нервная ткань характеризуется чрезвычайно медленным делением клеток, что приводит к тому, что межклеточный уровень отбора оказывается слабее внутриклеточного и с большой вероятностью может проявиться патогенный эффект, вызванный мутантными митохондриальными геномами.
Исследуя мутации в митохондриальной ДНК (мтДНК) нейронов мозга, две независимых лаборатории (Bender et al. 2006; Kraytsberg et al. 2006) получили интересные результаты о причинах старения и болезни Паркинсона - одной из основных нейродегенеративных болезней старости, характеризующейся нарушением планирования действий, что выражается в акинезии (ограничение произвольных движений), ригидности (повышение мышечного тонуса) и тремор конечностей (дрожание). Результаты их работы опубликованы в майском номере Nature Genetics .
Объектом исследований был головной мозг умерших людей, а именно участок под названием черное вещество ( substantia nigra ). Черное вещество находится в базальных ганглиях головного мозга (под корой больших полушарий), основная функция которых - регуляция произвольных движений: перехода от замысла (подготовки действия) к выполнению выбранной программы действия. При паркинсонизме происходит дегенерация нейронов черного вещества, выделяющих дофамин и регулирующих таким образом работу базальных ганглиев в целом. Конкретная причина дегенерации дофаминергических нейронов до сих пор была неизвестна.
|
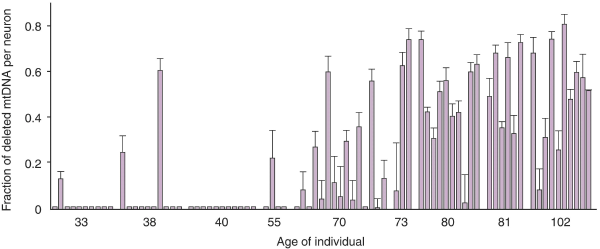
Техническим ноу-хау, позволившим сделать новые открытия, оказалась возможность исследования мутаций в митохондриальной ДНК (мтДНК) и доли мутантных геномов в каждом конкретном нейроне черного вещества, а не во всей нервной ткани.
Исследования нейронов черного вещества показали быстрое накопление делеций с возрастом. Оказалось также, что некоторые нейроны имеют сильный дефицит
|
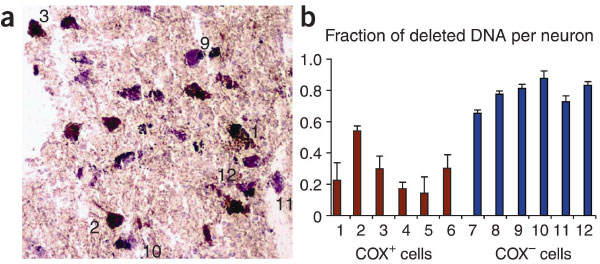
Важно отметить, что разные нейроны содержат уникальные делеции, что говорит о соматическом происхождении этих мутаций в нейронах. Интересно, что нейроны, выделенные из соседней области головного мозга (гиппокамп) старых индивидуумов не показывали высокого уровня делеций. Возможно, повышенная фоновая активность нейронов черного вещества сопряжена с образованием активных форм кислорода, которые, являясь мутагенами, приводят к образованию делеций в мтДНК. Активные формы кислорода могут появляться также и при метаболизме дофамина. Основываясь на этом предположении, для предотвращения появления делеций врачи предлагают обрабатывать нейроны черного вещества антиоксидантными препаратами.
Итак, соматические делеции в мтДНК играют важную роль в деградации нейронов при старении и болезни Паркинсона. Поскольку делеции появляются независимо в разных нейронах черного вещества примерно в одно и тоже время и затрагивают примерно одно и то же место, скорее всего существуют определенные
Другая потенциальная DILL-мутация может находиться в митохондрильной ДНК-полимеразе - ферменте, который реплицирует мтДНК. Мутантная форма данного фермента приводит к большому числу ошибок при репликации мтДНК, и линии мышей с таким ферментом страдают от раннего старения из-за накопления большого числа мутаций в мтДНК (Trifunovic et al . 2004).
Скорее всего, болезнь Паркинсона, как и старение, может быть вызвана многочисленными причинами. Например, мутация в ядерно-кодируемом гене альфа-синуклеине приводит к слипанию мутантных белков и образованию цитоплазматических включений, характерных для болезни Паркинсона (Polymeropoulos et al. 1997). Сходный эффект проявляется при нарушении у мышей автофагии - процесса удаления старых белков из цитоплазмы клеток (Hara et al. 2006).
Успех в изучении и лечении каждой из причин болезни Паркинсона и старения вносит свой вклад в общее дело борьбы с этим недугом. Впервые болезнь Паркинсона была описана в 1817 году Джеймсом Паркинсоном. Через век - в 1919 году - была предложена заместительная терапия посредством введения в кровь предшественника дофамина, что компенсирует функцию дегенерировавших нейронов черного вещества. В наше время научно доказана причина дегенерации нейронов и обсуждается возможность предотвращения появления этих делеций посредством антиоксидантов (Bender et al. 2006; Kraytsberg et al. 2006). Хочется верить, что к концу XXI века люди смогут приблизиться к причине еще ближе и научатся удалять DILL-мутации непосредственно из генома.
Источники:
1) Bender et al. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease // Nature genetics . 2006. V. 38. P. 515-517.
2) Kraytsberg et al. Mitochondrial DNA deletions are abundant and cause functional impairment in aged human substantia nigra neurons // Nature genetics . 2006. V. 38. P. 518-520.
3) G. Manfredi. mtDNA clock runs out for dopaminergic neurons // Nature genetics . 2006. V. 38. P. 507-508.
См. также:
В старении млекопитающих виноваты клеточные мутации, отвечающие за апоптоз, «Элементы», 15.07.2005.
Дополнительные источники:
G. A. Cortopassi. 2002. A neutral theory predicts multigenic aging and increased concentrations of deleterious mutations on the mitochondrial and y chromosomes (PDF, 49 Кб). Free radical biology & medicine . V. 33. P. 605-610.
E. A. Schon, R. Rizzuto, C. T. Moraes, H. Nakase, M. Zeviani, S. DiMauro. A direct repeat is a hotspot for large-scale deletion of human mitochondrial DNA. Science . 1989. V. 244. P. 346-349.
Trifunovic A., et al. 2004. Premature ageing in mice expressing defective mitochondrial DNA polymerase. Nature . V. 429. P. 417-423.
Taylor D.R., Zeyl C., Cooke E., 2002. Conflicting levels of selection in the accumulation of mitochondrial defects in Saccharomyces cerevisiae . PNAS. V. 99. P. 3690-3694.
Polymeropoulos M.H., et al. 1997. Mutation in the a-Synuclein Gene Identified in Families with Parkinson"s Disease. Science 276. P. 2045-2047.
T. Hara et al. 2006. Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature . Published online 19.04.2006.



 - Похоронный портал")
, First Woman to Record 24 Paganini Caprices - Похоронный портал")


